|
| |
СВЕТ И РАСТЕНИЯ: ФОТОСИНТЕЗ И ФИТООСВЕЩЕНИЕ
В статье даются основополагающие сведения о фотосинтезе - базисе существования и жизнедеятельности высших растений. Рассматривается в общих чертах внутренняя организация и функции листа и его светосинтезирующих единиц - хлоропластов, в т.ч. свойства входящих в их состав светочувствительных пигментов - хлорофиллов и каротиноидов, кратко описываются биохимические процессы и конечные продукты различных этапов фотосинтеза, а также приводятся современные данные о влиянии на фотосинтез спектрального состава и энергии освещения. Проблематика фотосинтеза рассматривается с привязкой к революционным изменениям в области источников искусственного фитоосвещения. Внимание! Статья настоятельно рекомендуется тем, кто собирается использовать фитолампы для выращивания рассады или для освещения теплиц и гроубоксов. Категория: общеобразовательная повышенного уровня.
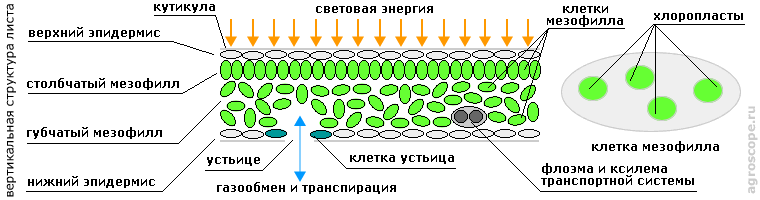
ВВЕДЕНИЕИз школьного курса ботаники известно, что фотосинтез является основой жизнедеятельности всех высших растений, и именно поэтому им нужно солнце. Благодаря фотосинтезу они производят из углекислого газа и воды необходимые вещества, являющиеся источниками энергии для всех биохимических процесов в их организме. Особенно популярной тема фотосинтеза стала в последнее время в связи с появлением светодиодных ламп (фитоламп), позволяющих качественно улучшить параметры искусственного освещения растений. Объем соответствующих публикаций зашкаливает, однако они в своей массе являются однобокими и зачастую недостаточно корректными. Цель данной статьи - дать растениеводам минимум знаний, который помог бы им сориентироваться в этой информационной лавине. ЛИСТ КАК ОРГАН ФОТОСИНТЕЗАОрганом фотосинтеза является лист, который одновременно выполняет и ряд других функций - дыхание, транспирацию (испарение) воды и синтез фитогормонов - гиббереллинов. Он имеет слоистую структуру: снаружи находится тонкий слой соединительной ткани - эпидермиса, защищенного слоем воскоподобного защитного вещества - кутикулой, а далее расположена основная ткань - мезофилл, которая обычно делится на плотную зону - столбчатый мезофилл, примыкающий к наружной светоприемной стороне листа, и рыхлую зону - губчатый мезофилл, в межклеточном пространстве которого циркулируют в процессе газообмена углекислый газ и кислород.
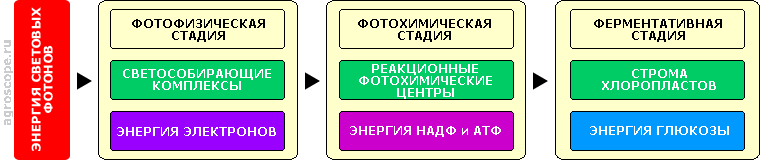
Структура листа как органа фотосинтеза Ткани мезофилла пронизаны сосудитстыми тканями транспортной системы, по которым циркулирует вода с растворенными питательными веществами, гормонами, ферментами и пр. На нижней (теневой) поверхности листа расположены устьица, через которые происходит газобмен и испарение воды. Устьица образованы двумя устьичными клетками, которые изменяют свою форму под воздействием определенных фитогормонов, открывая или закрывая таким образом устьичные отверстия. Клетки мезофилла содержат хлоропласты - автономные внутренние образования (органеллы), в которых и происходят все процессы, связанные с фотосинтезом. Их основу составляют молекулы фоточувствительных пигментов - хлорофилла A, хлорофилла B и каротиноидов. Основную массу данных пигментов составляют хлорофиллы, имеющие зеленые оттенки, именно они придают зеленый цвет листьям. Осенью хлорофиллы разрушаются и цвет листьев определяется цветом каротиноидов, имеющих желтые и оранжевые оттенки. Примечание. Такая структура тканей листа свойственна растениям т.н. С3 - типа, которые составляют большинство. Однако имеются еще и растения С4 и CAM типов, более характерные для тропических засушливых климатических зон, у которых структура тканей листа несколько иная. Это различие не затрагивает структуру фотосинтезирующих клеток и хлоропластов. Кроме хлорофиллов и каратиноидов в состав хлоропластов входят молекулы и других типов, участвующие в единой цепочке фотосинтеза, которая включает несколько стадий, выполняемых несколькими центрами.
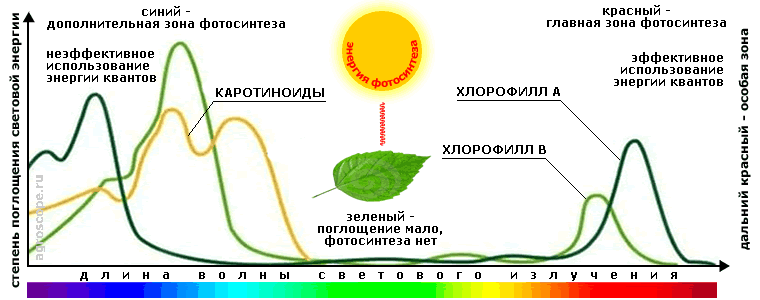
Структурная схема главной цепи фотосинтеза На первой стадии, именуемой фотофизической, энергия фотонов светового потока трансформируется в энергию орбитальных электронов молекул хлорофилла и каротиноидов, объединенных в светособирающие комплексы. На второй, фотохимической стадии накопленная энергия передается молекулам хлорофилла и белковых соединений фотохимических реакционных центров, стимулируя восстановление энергетического потенциала молекул НАДФ (носитель энергии для окислительно-восстановительных реакций) и синтез АТФ (носитель энергии для всех биохимических процессов). На третьей стадии - ферментативной, для которой свет не требуется, энергия НАДФ и АТФ используется во внутреннем пространстве хлоропластов (строме) для синтеза глюкозы из углекислого газа и воды в соответствии с уравнением: 6CO2 + 6H2O => C6H12O6 + 6O2 Синтезированная таким образом глюкоза поступает во внутриклеточное пространство, где претерпевает дальнейшие трансформации в процессе различных биохимических процессов - синтеза углеводов, белков, жиров, протохлорофиллов (полуфабрикатов для хлорофиллов), каратиноидов и пр. При этом необходимо отметить, что окончательный синтез хлорофиллов - их т.н. «зеленение», требует световой энергии, поэтому происходит на фотохимической стадии фотосинтеза. Для остальных же биохимических процессов свет как прямой источник энергии не требуется, однако он может быть необходим как источник информации для адаптации растения к условиям среды (см. ниже). Данная схема является весьма упрощенной, поскольку каждая из стадий имеет свою сложную внутреннюю организацию со множеством существенных параметров и зависящую от множества внешних факторов. Тем не менее, она дает верное представление о трансформации световой энергии и совокупности процессов фотосинтеза, что важно для понимания их принципиальной сути. СПЕКТРАЛЬНЫЕ ПАРАМЕТРЫ ФОТОСИНТЕЗАСпектральная характеристика фотосинтеза это именно тот параметр, который вышел на передний план в связи с появлением светодиодных источников света, позволяющих в определенных пределах видоизменять спектр излучения. Практически все публикации, касающиеся искусственного освещения растений, сегодня начинаются с соответствующего «ликбеза», часто весьма поверхностного и некорректного. Поэтому вопрос соотношения спектра освещения и параметров фотосинтеза требует особо пристального рассмотрения. Спектр поглощения листа Падающая на листья световая энергия, если пренебречь их светопропусканием, делится на отраженную и поглощенную. Отражается энергия в зеленой области спектра, в соответствии с цветом листьев. В этой области, в связи с почти полным отстутствием поглощения энергии, фотосинтез не идет. Основной же вклад в энергию фотосинтеза вносит поглощение световой энергии хлорофиллами A и B в красной области, при этом несколько бОльшую (в 1.2 -1.4 раза) часть вносит хлорофилл А, что обусловлено преобладанием его молекул в хлоропластах. Максимумы поглощения хлорофиллов сдвинуты относительно друг друга, что расширяет спектральный диапазон поглощения и в определенной степени выравнивает производительность фотосинтеза при суточном изменении спектра.
Спектр фотосинтеза На прямом солнечном свету энергии красного диапазона спектра вполне достаточно для удовлетоврения потребностей растения в продуктах фотосинтеза. Однако, если прямого солнечного света по каким-либо причинам (препятствия, облачность) не хватает, то «выручает» энергия, полученная в синем диапазоне, который характерен для рассеянного света зенитальной области небесной сферы, при этом преобладающая доля этой энергии поглащается каротиноидами. Эффективность же «усвоения» процессами фотосинтеза энергии синего диапазона существенно ниже, чем красного, несмотря на несколько больший коэффициент его поглощения растениями. Это обусловлено особенностями квантовой природы света и физики процессов в зеленом листе, которые требуют отдельного рассмотрения. Особенности численных показателей спектра фотосинтеза В популярных публикациях часто оперируют конкретными значениями длин волн спектра фотосинтеза. Для того, чтобы делать какие-то конкретные выводы из такого рода информации надо иметь в виду следующее:
Положение еще осложняется тем, что указанные особенности могут быть различными для разных растений. При этом также следует учитывать, что пигменты выстраиваются в цепочки передачи энергии возбуждения, образуя две фотосистемы, у которых длины волн поглощения последних молекул в цепочках реакционных центров равны 700 нм (фотосистема 1) и 680 нм (фотосистема 2). Результатом является то, что избыточная энергия квантов более коротких длин волн при прохождении цепочки уходит в потери. Как видно, математика спектра фотосинтеза весьма сложна и зависит от многих обстоятельств, поэтому ко всевозможным популистким выводам, основанным на манипуляции с какими-либо численными величинами спектров поглощения, что встречается достаточно часто, надо относиться с крайней осторожностью.
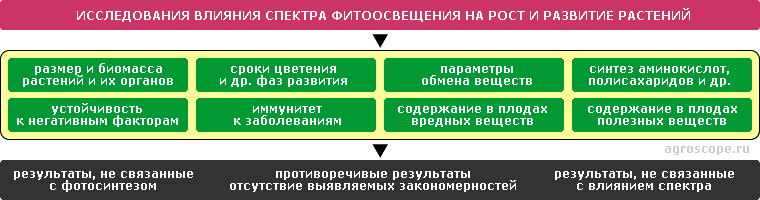
К вопросу о численных значениях спектров поглощения фотосистем Влияние спектра на рост и развитие растений Появление широких возможностей по регулированию спектра искусственного освещения породило желание использовать его для управления какими-либо параметрами выращиваемых культур аналогично тому, как это делатеся с помощью агропрепаратов - регуляторов роста. Однако современная теория фотосинтеза не допускает таких возможностей, поскольку, как было сказанно выше, ограничивает продукцию фотосинтеза только глюкозой и хлорофиллами Тем не менее, соответствующие исследования ведутся в надежде открыть новые механизмы. В частности, в ряде публикаций утверждается об обнаружении влияния спектра освещения на рост и биомассу растений и их органов, на синтез аминокислот, полисахаридов и пр., на содержание нитратов и полезных веществ, на устойчивость к стрессам и иммунитет и т.д. Однако эти данные не носят закономерного характера и часто противоречивы, что напоминает результаты исследований, связанных с лунным фактором. При ближайшем рассмотрении видно, что часть их не имеет отношения к фотосинтезу, а может быть связана с сигнальной функцией света - фотоморфогенезом, а часть объясняется изменением суммарной энергии различных спектральных областей. Подробный анализ такого рода исследований не входит в задачи настоящей статьи и является предметом отдельных аналитических работ.
Исследования влияния спектра на рост и развитие растений О ЕДИНИЦАХ ИЗМЕРЕНИЯ ФИТООСВЕЩЕНИЯВ связи с описанными здесь особенностями поглощения и использования растениями световой энергии, а именно в связи с зависимостью от длины волны и квантовым характером процессов, стандартные световые физические единицы мало пригодны для оценки световых параметров фитоламп и величины создаваемой ими освещенности, в связи с чем для этих целей были введены специальные единицы:
Однако эти величины также несовершенны, т.к. учитывают только крайние границы диапазона фотосинтеза и его квантовый механизм, но не учитывают неравномерности использования световой энергии растениями в различных участках спектра. В связи с этим для более точных оценок используется более совершенная единица - усваиваемый растением поток фотонов (YPF - yield photon flux), которая учитывает типовой спектр фотосинтеза. Оценка же эффективности фитоламп для конкретных культур требует знания полных спектральных характеристик как самих культур, так и фитоламп, что является нетривиальной задачей.
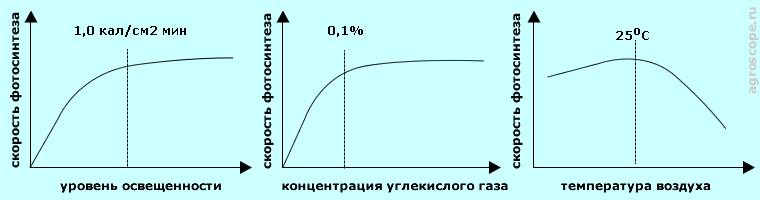
К вопросу о единицах измерения фитоосвещения ВЛИЯНИЕ НА ФОТОСИНТЕЗ УРОВНЯ ОСВЕЩЕННОСТИ И ПАРАМЕТРОВ СРЕДЫДля нормального течения процесса фотосинтеза нужны, кроме требуемого спектра, требуемый уровень освещенности в заданном спектре, требуемая концентрация углекислого газа, оптимальный температурный диапазон и нормальный водный режим. Зависимость скорости фотосинтеза от уровня освещенности носит логарифмический характер, т.е. на начальном участке она растет по закону, близкому к линейному, затем рост ее замедляется и, в конечном итоге, она достигает насыщения. Это связано с тем, что начиная с определенной величины интенсивности фотосинтеза лимитирующими становятся не световые реакции, а скорости газообмена и темновых реакций. Для разных растений порог насыщения по уровню освещенности различен и по его значению различают светолюбивые, теневыносливые и тенелюбивые культуры.
Влияние на фотосинтез уровня освещенности и параметров среды Зависимость скорости фотосинтеза от содержания в воздухе углекислого газа имеет аналогичный характер. Насыщение наступает при концентрациях 0,06% - 0,15%, что превышает концентрацию углекислого газа в атмосфере, равную в среднем 0,04%. Данное свойство используют для ускорения тепличного оборота и повышения урожайности некоторых культур, в частности, огурцов, искусственно повышая концентрацию углекислого газа в теплицах. А вот зависимость от температуры воздуха несколько иная - сначала вплоть до температуры 25°C - 30°C отмечается незначительный линейный рост, а затем начинается резкий спад. Соответственно в теплицах и парниках необходимо поддерживать не максимальную, а оптимальную температуру. Водный режим косвенным образом воздействует на газообмен. При нормальном снабжениии растения водой устьица открыты полностью и поступление углекислого газа не ограничивается. При дефиците воды в целях ее экономии растение уменьшает объем транспирации, прикрывая устьица, соответственно ограничивается поступление углексилого газа, в результате чего снижается скорость фотосинтеза. Все сказанное относится к растениям С3 - типа. У растений C4 - типа более высокая производительность фотосинтеза и, соответственно, более высокие пороги насыщения по уровню освещенности и температуре воздуха. КРАТКОЕ РЕЗЮМЕФотосинтез является сложным физико-химическим процессом, имеющим квантовую природу и зависящим от спектра и интенсивности освещения, а также от концентрации углекислого газа, температуры воздуха и водного режима. Оптимизация данных параметров способна повысить производительность фотосинтеза и, соответственно, сократить сроки плодоношения и повысить урожайность садово-огородных культур. Появившиеся на рынке светодиодные источники позволяют повысить экономичность фитосветильников и формировать заданный спектр искусственного освещения растений. Однако возможность достижения регулированием спектра многих рекламируемые эффектов сомнительна, поскольку не имеет под собой ни общепризнанной теоретической базы, ни надежного экспериментального подтверждения. В такой ситуации применение дорогостоящих светодиодных фитоламп для достижения обещаемых их производителями преимуществ сопряжено с определенным риском и требует обдуманного выбора, основанного на знании основ фотосинтеза.
Свет и растения: фотосинтез и фитоосвещение |
# БИБЛИОТЕКА #
| |
|
| |
|